")
")
Нерпа
Уникальной особенностью пресноводной экосистемы Байкала является наличие в ней эндемичного байкальского тюленя - нерпы (Phoca sibirica), высшего звена пищевой цепи. В последние годы биология байкальской нерпы очень хорошо изучена, в особенности благодаря работам В.Д. Пастухова (1993) и его учеников. Специально для Байкала им разработана оригинальная методика учета тюленей (Пастухов, 1993, с. 214). Нерпа рождает и выкармливает своих детенышей зимой, когда Байкал покрыт льдом. Для этой цели она строит из снега логово вблизи той лунки, в которую ныряет для добычи корма. В логове живет щенок. Новорожденные щенки нерпы, в отличие от темно окрашенных взрослых животных, имеют светло окрашенный мех. Весной щенок линяет, оставляя вылинявшую шерсть в логове. По мере наступления весны снег тает, и логово разрушается, а на его месте остается вылинявшая шерсть.
Методика учета по В.Д. Пастухову (1993), внедренная им в 1972 г., состоит в том, что исследователи, двигаясь по всему Байкалу от юга к северу на мотоциклах по тающему льду, обследуют поперечные разрезы через акваторию озера. Число таких разрезов составляло от 12-14 в 70-е гг. до 27 в 1994 г. На каждом разрезе они останавливаются на нескольких определенных точках и вокруг этих точек на площади 1,5 км подсчитывают разрушенные логова, возле которых найдена вылинявшая светлая шерсть. Экстраполируя полученные результаты по специальной методике на всю площадь Байкала, определяют число щенков, родившихся в год учета, а следовательно и число оплодотворенных ранее самок. Зная эту величину и показатели возрастно-половой структуры популяции (найденные путем исследования материала, полученного охотниками при добыче нерпы), а также некоторые дополнительные данные, в конечном итоге можно рассчитать общую численность байкальской нерпы.
Нужно отметить, что учет популяции диких млекопитающих очень сложен и обычно не обеспечивает высокую точность, давая лишь порядковую оценку, которая может отличаться от истинной численности в несколько раз. Для учета байкальской нерпы до появления методики В.Д. Пастухова неоднократно предпринимались попытки использовать авиационные и другие методы, но они не дали достоверных результатов. Методика же В.Д. Пастухова имеет высокую точность. Можно предполагать, что при уровне достоверности 0,95 эта точность составляет 20-30%.
На рис. 3.3.1 показана динамика численности байкальской нерпы в период с 1950 по 1994 гг. по данным разных авторов; эти данные заимствованы нами из краткого обзора Е.А. Петрова и др. (1997а). Численность, найденная в 1950 - 1960-е гг., до создания методики В.Д. Пастухова, по-видимому, была существенно занижена. Численность, определенная по новой методике после 1972 г., варьирует в пределах 70 - 140 тыс. животных. Различия данных последних лет, полученных Е.А. Петровым в 1988 и 1994 гг., и В.Д. Пастуховым в 1973 - 1985 гг., вероятно, вызваны не ростом популяции, а введенными небольшими изменениями методики учета.
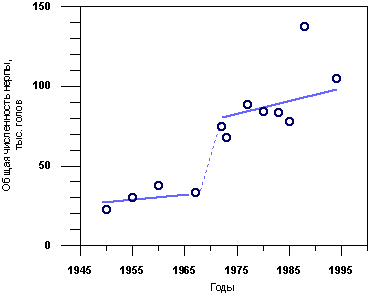
Рис. 3.3.1. Численность байкальской нерпы по данным разных авторов. Данные из обзора Петрова и др. (1997 а). Методика ледового учета внедрена В.Д.Пастуховым в 1972 г. Учет в 1988 и 1994 гг. проведен под руководством Е.А. Петрова.
![]() Загрузить исходные данные в формате Grapher 3
Загрузить исходные данные в формате Grapher 3
Согласно данным последних лет, существенно не изменилась не только численность нерпы, но и возрастно-половая структура популяции (Петров и др., 1997а, 1997б; Петров, 1997; Петров, Егорова, 1998). Данные учета в пределах их точности не выявили изменений, которые могли бы быть вызваны накоплением хлорорганических соединений в жире нерпы (см. раздел 2.3). таким образом, полученные данные не дают никаких оснований предполагать изменения популяции байкальской нерпы под действием антропогенных нагрузок.
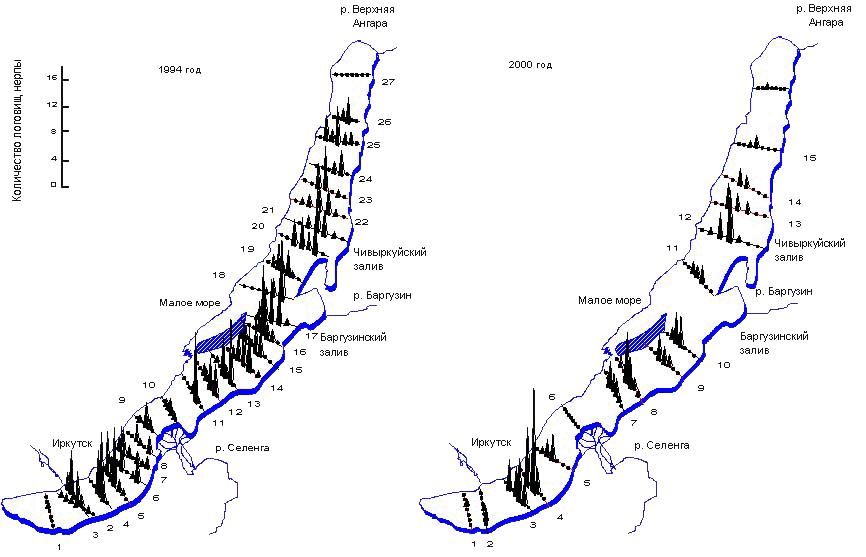
Рис. 3.3.2. Данные учета логовищ нерпы
В 2000 г. учет популяции байкальской нерпы был выполнен под эгидой международной организации Гринпис. Его результаты сформулированы в отчете (Пастухов, 2001), который (иногда вопреки его содержанию) послужил поводом для широковещательных заявлений о том, что произошло существенное снижение численности байкальской нерпы, в особенности в северной части озера.Учет был выполнен по методике В. Д. Пастухова, но на недостаточном числе разрезов. О том, что точность измерений не позволяет сделать однозначный вывод о падении численности нерпы, наглядно свидетельствуют данные, представленные на рис. 3.3.2. Во время учета 2000 г. были пропущены важные разрезы.
Численность приплода в 1994 г. составила 21 933 ± 3 960 голов нерпы. Было заложено 27 разрезов, обследовано 183 площадки общей площадью 412 км2. В 2000 г. численность приплода составила 17862 ± 5848 голов нерпы. Было заложено 16 разрядов, обследовано 112 площадок общей площадью 252 км2.
Очевидно, что методике ледового учета байкальской нерпы следует искать альтернативу, так как она весьма трудоемка, опасна для исполнителей и недостаточно точна (прежде всего вследствие неравномерности распределения зверя по акватории Байкала) для принятия быстрых решений о режиме хозяйственного изъятия. Перспективными с этой точки зрения представляются акустические методы, но для тюленей они пока в мировой науке не разработаны.
Большое внимание общественности к благополучию популяции байкальской нерпы было привлечено осенью 1987 - зимой 1988 гг., так как в это время произошла массовая гибель этих животных. Заболевшие нерпы с характерными симптомами (малая подвижность, паралич конечностей, конъюнктивит) выбрасывались на берег и там погибали. Возникли подозрения, что гибель нерпы вызвана поступившими в Байкал неизвестными ядовитыми веществами. Учет трупов погибших животных, проведенный весной и летом 1988 г., показал, что в общей сложности погибло около 6 тыс. тюленей.
Некоторые особенности динамики распространения заболевания по акватории Байкала и клинические признаки позволили предположить, что причиной гибели животных было инфекционное заболевание, в чем-то похожее на собачью чумку. Применение методов молекулярной биологии уже в феврале 1988 г., то есть спустя три месяца после начала эпизоотии, позволило однозначно установить, что причиной массовой гибели является мoрбилливирус, аналогичный вирусу собачьей чумки и родственный вирусу кори человека (Grachev et al., 1989; Likhoshway et al., 1989; Вспышка.., 1992).
Интересно отметить, что в мае 1988 г., через полгода после начала байкальской эпизоотии стала наблюдаться массовая гибель тюленей, живущих в Балтийском и Северном морях. Вскоре было установлено, что и эта болезнь вызвана морбилливирусом (Osterhaus et al., 1988). Впоследствии оказалось, что европейский вирус сильно отличается от байкальского, и никакой прямой связи между этими эпизоотиями нет. Не подтвердилось и предположение и о том, что агентом, вызвавшим болезнь байкальских и европейских тюленей, была живая вакцина, широко применяющаяся для защиты домашних животных - собак, норок и хорьков (Mamaev et al., 1995). Оказалось, что морбилливирус вызывает системное поражение многих органов и тканей тюленей (Belykh et al., 1997).
Вирусы, вызвавшие эпизоотии на Байкале и в Западной Европе, а также пораженные ими популяции тюленей стали предметом большого числа научных публикаций ученых из России, Голландии и Великобритании. В России этими работами руководил недавно умерший Л.В. Мамаев. На рис. 3.3.3 приведены последовательности нуклеотидов фрагментов генов фосфобелка некоторых морбилливирусов (Mamaev et al., 1996), а именно вакцинного штамма Onderstepoort (CDV Ond); изолята вируса собачьей чумки, полученного от больной собаки (CDV dog); изолята морбилливируса европейских тюленей (PDV-1 Eur); изолята морбилливируса байкальской нерпы (PDV-2 Bai). Кроме того, приведена последовательность нуклеотидов того же фрагмента, найденная путем расшифровки структуры РНК, выделенной из мозга больной байкальской нерпы (PDV-2 RNA). Можно видеть, что байкальский вирус наиболее близкородствен вирусу собачьей чумки. Вирус продолжает циркулировать в популяции байкальской нерпы по настоящее время, не вызывая ее массовой гибели, поскольку большинство животных содержит в крови высокие титры антител против вируса собачьей чумки (Н. Деникина, Т. Бутина, частное сообщение).
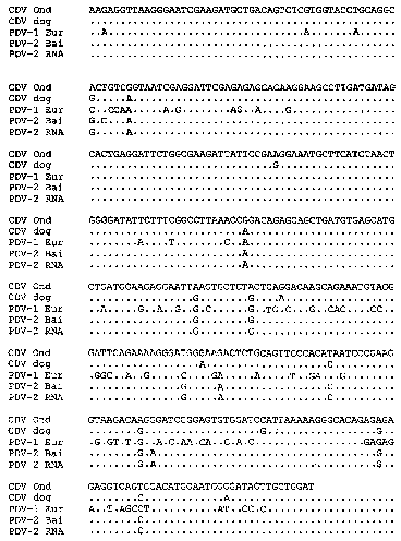
Рис. З.З.З. Последовательности нуклеотидов фрагментов гена фосфобелка пяти морбилливирусов. CDV Опа-вакцинный штамм Onderstepoort; CDV dog-изолят от собаки, больной чумкой: PDV-1 Eur-изолят от европейских тюленей; PDV-2 Bai-изолят от байкальских тюленей; PDV-2 RNA-продукт амплификации вирусной РНК из мозга байкальской нерпы (Mamaev etal., 1996).
В литературе иногда высказываются предположения, что байкальские и европейские тюлени были поражены морбилливирусами в 1987-1988 гг. по причине ослабления их иммунитета из-за накопления высоких концентраций хлорорганических веществ (см. например, Nakata et al., 1997). Такое предположение имеет право на существование, поскольку известно, что хлорорганические вещества могут воздействовать на некоторые тонкие механизмы иммунного ответа млекопитающих. Однако убедительных доказательств взаимосвязи морбилливирусных эпизоотий с накоплением хлорорганических веществ в тканях животных до настоящего времени не получено. Вряд ли можно предполагать, что иммунная система байкальской нерпы существенно нарушена, поскольку в крови этих животных и в период их массовой гибели и позднее имелся высокий титр противовирусных антител, а эпизоотия прекратилась, хотя уровень химического загрязнения не изменился.
Скорее всего, эпизоотии среди тюленей Байкала и Западной Европы возникли независимо друг от друга по чисто природным причинам. Например, исследования крови европейских тюленей, собранной до эпизоотий 1988 г. и хранившейся в замороженном состоянии, показали, что еще до эпизоотии тюлени имели контакт с вирусом, весьма близко родственным тому, который поразил их в 1988 г. Некоторые исследователи предполагают (см. Mamaev et al., 1995), что морбилливирусы циркулируют в популяциях тюленей Европы и Северной Америки постоянно, со временем несколько видоизменяясь вследствие мутаций. К вспышке заболевания мог привести контакт изолированных ранее популяций, являющихся носителями разных штаммов вируса. Такой контакт, например, мог иметь место в 1986 -1987 гг., когда произошла массовая миграция арктических тюленей из Баренцева моря в моря Северной Европы (см. Mamaev et al., 1995).
По Байкалу исторических сведений о более ранних вспышках морбилливирусной инфекции нет. Данные молекулярно-биологических исследований позволяют считать вполне вероятным, что источником вируса эпизоотии 1988 г. были собаки или другие хищные животные. Вирус собачьей чумки (чумы плотоядных) циркулирует в популяциях хищников постоянно и мог попасть в Байкал в любое время. То обстоятельство, что вирусное заболевание у тюленей было зарегистрировано в 1987-1988 гг. впервые, может быть объяснено не тем, что нерпа раньше чумкой не болела, а тем, что предыдущие вспышки имели место давно, когда эта популяция еще не привлекала нужного внимания, а нерпа рассматривалась как вредное животное, снижающее уловы рыбы.
Известно, что морбилливирусная инфекция поражает малые популяции периодически, и что перерывы между вспышками могут быть весьма длительными (Клифф, Хаггет, 1984). Как видно из рис. 3.3.4, до введения массовой вакцинации в 1964 г., вирус кори, относящийся к группе морбилливирусов, поражал население США ежегодно. В этот же период с 1945 до 1965 гг. в малонаселенной Исландии вспышки кори происходили с интервалами в 3-4 года. До 1945 г., когда контакты Исландии с внешним миром были менее регулярными, перерывы между вспышками достигали 10 и более лет. Следовательно, можно предполагать, что следующая вспышка заболевания байкальских тюленей собачьей чумкой вряд ли возникнет скоро, так как процент защищенных антителами животных в ней весьма высок, а новорожденные тюлени получают антитела с материнским молоком, и, если даже они заболевают, то болезнь протекает в легкой форме.
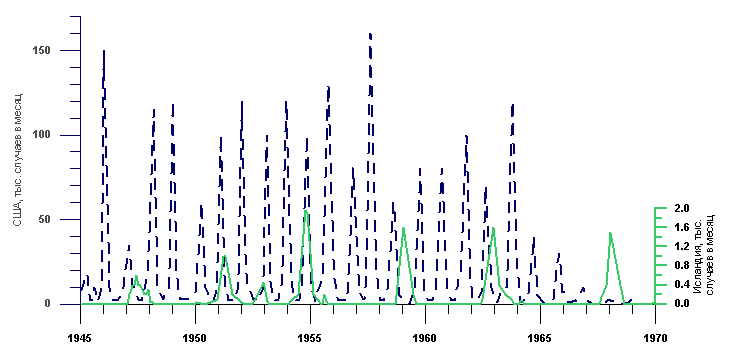
Рис. 3.3.4 Динамика заболеваемости корью в США (население 200 млн чел.) и в Исландии (население 200 тыс. чел.). Клифф, Хаггет, 1984.
![]() Загрузить исходные данные в формате Grapher 3
Загрузить исходные данные в формате Grapher 3
Просмотреть список литературы (формат PDF)
Перепечатка без согласия автора запрещена. E-mail для контактов: [email protected]
Публикация книги в сети интернет выполнена по гранту РФФИ-байкал №05-07-97200 E-mail: [email protected]